IUCN/SSC Otter Specialist Group Bulletin

|
< ©IUCN/SCC Otter Specialist Group Volume 28A Proceedings Xth International Otter Colloquium, Hwacheon, South Korea Citation: Hájková, P., Zemanová, B., Roche, K. and Hájek, B. (2011) Conservation Genetics and Non-Invasive Genetic Sampling of Eurasian Otters (Lutra lutra) in the Czech and Slovak Republics . Proceedings of Xth International Otter Colloquium, IUCN Otter Spec. Group Bull. 28A: 127 - 138 Conservation Genetics and Non-Invasive Genetic Sampling of Eurasian Otters (Lutra lutra) in the Czech and Slovak Republics Petra Hájková1, Barbora Zemanová1, Kevin Roche1 and Bedřich Hájek2
1Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic, Květná 8, 603 65 Brno, Czech Republic, e-mail: hajkova@ivb.czhajkova@ivb.cz |
    |
| (Received 23rd March 2011, accepted 27th December 2011) |
|
Abstract: Many important basic parameters for successful conservation management of Eurasian otter (Lutra lutra) populations are still not fully understood. Otters are rare and elusive animals, and it is very difficult to study their abundance and population structure. Over the past years, however, genetic methods have provided a new approach to this problem. If non-invasively obtained samples, such as faeces, are analysed, disturbing or even observing the animals is not required. We used a non-invasive genetic method to assess population size, sex ratio and spatial distribution of otters in two different habitats. Further, we studied the genetic structure of otters in the Czech and Slovak Republics using DNA extracted from both tissue and spraint samples. A total of 448 spraint samples and 134 tissues were analysed using 10 microsatellite loci and a SRY marker. At the first study site, a mountainous area of Slovenský raj National Park, 13 individuals (7 males and 6 females) were identified, while at the second site, a fishpond area of Třeboňsko Biosphere Reserve, it was 50 individuals (29 males and 21 females) for the same area size (100 km2). When studying genetic structure of otter populations in the Czech and Slovak Republics, a relatively high level of genetic differentiation was found between the Czech and Slovak populations (FST = 0.154), supported by a perfect assignment in Bayesian clustering. Both MSVAR and BOTTLENECK programmes, which infer demographic history, indicated the occurrence of recent population bottleneck in both populations. Despite recent recovery, otter populations in both countries are still vulnerable. |
| Keywords: microsatellites, spraints, abundance, sex ratio, genetic structure, bottleneck, effective population size, conservation management |
| Française | Español |
INTRODUCTION
Over the latter part of the 20th century, Eurasian otter (Lutra lutra) populations suffered dramatic declines in many European countries (Macdonald and Mason, 1994). In the Czech Republic, most of the otter population became totally isolated around the 1970-80s, while the Slovak population remained partly connected to the relatively continuous Central and Eastern European otter distribution range. Changes in habitat quality and food availability, chemical pollution and direct persecution have resulted in the extinction of the otter over much of its previous territory, and fragmentation of remaining populations. These events may have led to a loss of genetic variability, a decrease in fitness, and increased genetic differentiation among populations, which are the factors that may affect populations’ viability (Frankham et al., 2002).
Recently, otter populations have started to recover in most European countries (e.g. Conroy and Chanin, 2002; Mason and Macdonald, 2004). Otter numbers are increasing and many populations that had become separated over the last century are now, once again, rejoining. However, as the range of the otter increases, the historic conflicts between fish-farmers/anglers and the otter are once again emerging, particularly in the Czech Republic, and environmental and conservation bodies are facing increasing pressure for a reduction in otter numbers through legal culls. However, because the Czech and Slovak otter populations experienced a population decline of unknown extent over the past century, their genetic polymorphism and effective population size may have been depleted, making them vulnerable to any violent demographic change. It is crucial, therefore, that the genetic variability and structure of the extant populations is assessed, in order to help to provide scientifically informed recommendations for their future conservation management.
The Eurasian otter is an elusive, nocturnal animal that is rarely observed and therefore difficult to study. Consequently, many important ecological parameters that are essential for their successful conservation management are still not fully understood. Significant problems arise with the need for reliable assessment of basic population parameters such as abundance and population structure. Over the past years, however, genetic methods have become available also for such studies. If non-invasively obtained samples, such as faeces, hair, urine, etc. are analysed, disturbing or observing the animals is not required. For elusive mammals such as otters, spraints often represent the only available biological material. Spraints contain cells shed from the gut lining, and the DNA can potentially be extracted and analysed. Genetic methods are based on typing of several microsatellite (highly variable simple sequence repeats) and SRY (a male specific gene located on the Y chromosome) markers, and can potentially provide identification of individuals, sex, relatedness, estimates of population size and the level of genetic polymorphism within or between populations (Kohn and Wayne, 1997).
Our project had two aims: (1) to assess population size, sex ratio and spatial distribution of otters in two different habitats using a non-invasive genetic sampling; and (2) to estimate the genetic variability and population genetic structure of otters in the Czech and Slovak Republics using microsatellites.
ANIMALS, MATERIAL AND METHODS
Samples and molecular analyses
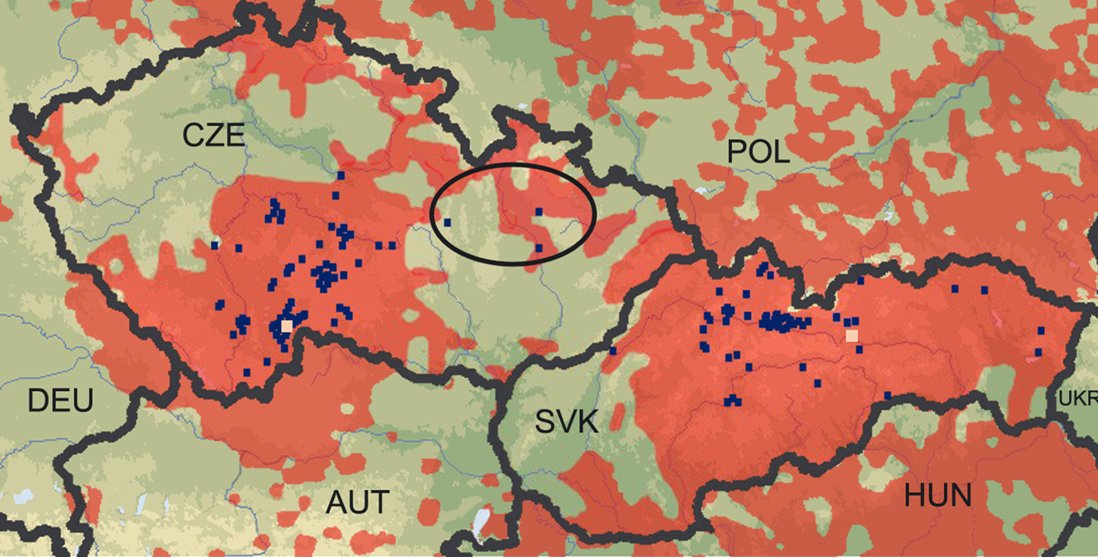
Two sample types were used in this study, tissue samples and faeces (spraints). Tissue samples (muscle) were collected from 134 otter carcasses from different locations in the Czech (n = 82) and Slovak (n = 52) Republic (Fig. 1). The samples were collected between 2002 and 2005, and were preserved in 96% ethanol and stored at -20 °C until DNA extraction. Spraint samples originated from two study sites of 100 km2 (Fig. 1); the first located in South Bohemia (Třeboňsko Biosphere Reserve & Protected Landscape Area), and the second in Eastern Slovakia (Slovenský raj National Park). Fresh spraints were collected in cold months between 2003 and 2004, preserved in 96% ethanol or different kit buffers (see Hájková et al., 2006), and stored at -20 °C or -80 °C. For DNA extraction methods and polymerase chain reaction (PCR) conditions see Hájková et al. (2006) and Hájková et al. (2007). Ten microsatellite loci were assayed, i.e. Lut701, Lut715, Lut717, Lut733, Lut782, Lut818, Lut832, Lut833, Lut902, Lut914, and Lut-SRY marker (Dallas and Piertney, 1998; Dallas et al., 1999; Dallas et al., 2000). PCR products were electrophoresed on ABI Prism 310 and 3130 Genetic Analysers and analysed using GeneScan 3.7 or GeneMapper 3.7 software (Applied Biosystems).
 |
|
Figure 1. Present distribution (red) of the Eurasian otter in the Czech and Slovak Republic and surrounding countries (at the time of collection of the samples), with sites marked where material for genetic analysis was obtained (blue squares = carcasses; beige squares = sites of non-invasive genetic sampling; ellipse = area of reintroduction). Otter distribution map adapted from Urban and Kadlečík, 2001; Reuther and Krekemeyer, 2004; Roche et al., 2004. Click for larger image |
PCR amplification for tissue DNA was repeated two to four times according the quality and quantity of extracted DNA, as some of the samples were obtained from highly degraded carcasses. For spraint DNA, the multiple-tubes approach was followed (Taberlet et al., 1996), with a minimum of six positive PCR for homozygotes and three for heterozygotes; however, up to 12-16 positive PCRs were required in some cases to confirm the genotype in samples with high level of allelic dropout. Stringent precautions were taken to avoid contamination when working with spraint DNA.
Study areas of non-invasive genetic sampling (NGS)
Two study areas, each of 100 km2, were selected. The first one was a mountainous/sub-mountainous area located in the Slovenský raj National Park and its surroundings in Slovakia (hereafter ‘Slovenský raj’, or ‘SR’); the second one was a fishpond region located in the Třeboňsko Protected Landscape Area & Biosphere Reserve in the Czech Republic (hereafter ‘Třeboňsko’, or ‘TR’). The Slovenský raj site consisted of an oligotrophic river and its main tributaries (two streams), with a total length of watercourses of ca. 50 km. The Třeboňsko site included a lowland river and a high number of mostly large eutrophic fishponds, connected by a complex system of channels. Both sites were known to support resident otter populations, and were parts of the relatively large and continuous populations of wider surroundings. For methodological reasons, we kept both study areas at the same size, despite different otter numbers could be expected in such different habitats (see also Hájková et al., 2009).
Data analyses
For non-invasive samples, software GEMINI (Valière et al., 2002) was used to calculate probability of identity (PI), i.e. the probability that two individuals drawn at random from a population will have identical multi-locus genotypes. Both ‘random PI’, i.e. theoretical expected PI based on allele frequencies from non-invasive genotypes obtained (herein PI), and PI for siblings, which is a conservative upper bound for the probability of identity (herein PIsibs) were calculated (Paetkau and Strobeck, 1994; Taberlet and Luikart, 1999; Waits et al., 2001). Frequency of allelic dropout (ADO) was computed as the ratio of the number of observed ADO at given locus on the number of positive amplifications of individuals determined as heterozygous according to their consensus genotype at this locus (Broquet and Petit, 2004). Unspecific amplification, including polymerase slippage artefacts and occasional contamination, were counted as false alleles (FA).
For population genetic analyses following programmes were used: FSTAT (Goudet, 2001), GeneClass2 (Piry et al., 2004), GENETIX (Belkhir et al., 1996-2004), and STRUCTURE (Pritchard et al., 2000). Recent population bottlenecks were inferred using BOTTLENECK (Cornuet and Luikart, 1996), and both recent and past demographic history of populations was inferred using MSVAR (Beaumont, 1999; Storz and Beaumont, 2002). MSVAR programme was used to estimate effective population size (Ne; Frankham et al., 2002; Mills, 2007). More details on methods can be found in Hájková et al. (2007) and Hájková et al. (2009).
RESULTS
Non-invasive genetic sampling
A total of 198 spraint samples were collected from the Slovenský raj site and 250 from the Třeboňsko site, respectively. Most of the samples were characterised as spraints, followed by anal jelly samples. Approximately 60% of the samples were successfully genotyped; 63% (125 samples) from SR site and 55% (137 samples) from TR site, respectively. The highest success rate, 82%, was obtained for anal jelly samples, followed by 58% in case of spraints with jelly, and the lowest, 46% for spraint samples. For a complete set of 10 loci, PI was 5.846 × 10-6 and PIsibs 3.665 × 10-3 for samples from the Slovenský raj site; and 1.211 × 10-5 and 5.144 × 10-3 for samples from the Třeboňsko site, respectively. The mean frequency of allelic dropout was 17.9% (SD = 0.275), but its single values for particular samples and loci varied greatly, from 0% to 100%. False alleles were recorded on the average in 2.9% of samples (SD = 0.123). Average number of positive PCRs per locus and sample was 4.6 (range 2-16).
At the SR site, successfully genotyped samples yielded 13 different genotypes, i.e. individuals SR1 - SR13. Seven individuals were males and six females (sex ratio 1.2:1). Otter density for this site was 0.13 individual per 1 km2; or, more meaningfully (considering linearity of the habitat), one individual per 3.8 km of watercourses. The number of positive samples per individual ranged from 1 to 23 (mean 9.6, SD = 7.32). Two individuals were identified based on a single sample. Seven individuals were assigned as residents, being recorded repeatedly in the time span of more than one month. Their minimum home range size (linear pattern) ranged between 2 and 15 km in males, while in females between 3 and 8 km.
From TR site, 50 different genotypes were obtained, corresponding to individuals TR1 - TR50. Thus otter density in this area was 0.5 individual per 1 km2. From 50 individuals identified, 29 were males and 21 females, resulting in the sex ratio 1.4:1. Number of positive samples per individual ranged from 1 to 13 (mean 2.8, SD = 2.58), but approximately half of the otters, 26, were identified from a single sample. Altogether 13 individuals were assigned as residents, being recorded repeatedly in the time span of more than one month. For the residents, minimum home range size was assessed as the distance between the most distant points of their occurrence. In males, it ranged from 600 to 4600 m, while in females from 200 to 1000 m.
Genetic variability and population genetic structure
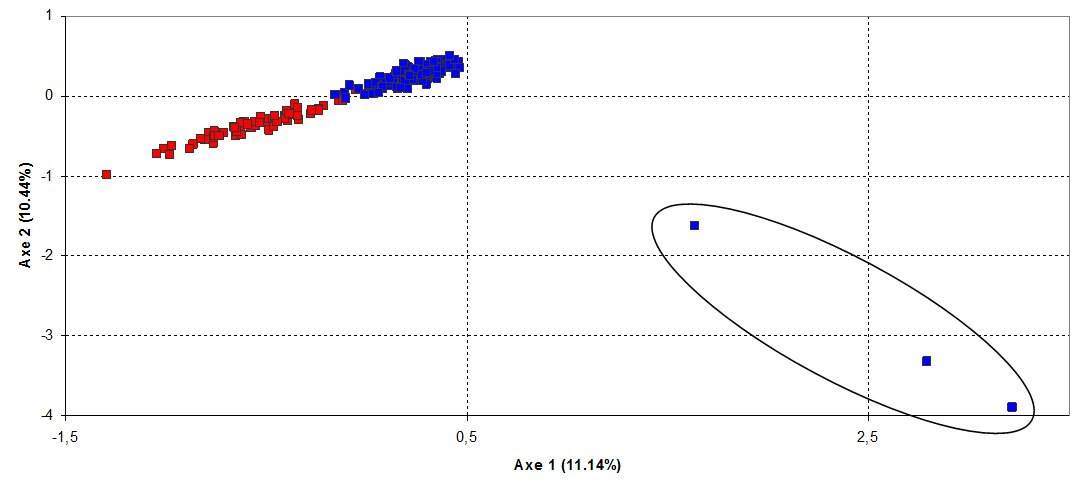
Genotypes for 10 microsatellite loci were obtained for 132 otters from the Czech Republic and 65 from the Slovak Republic, respectively (Fig. 1). All analysed loci were polymorphic in both samples, ranging from 3 to 6 alleles. Characteristics of genetic variability (e.g. allelic richness, observed and expected heterozygosity) were moderately high (Table 1). Their values were slightly higher in Slovak population, but differences were not statistically significant. The Czech otter population was in Hardy-Weinberg equilibrium (HWE; FIS = 0.023, P=0.143) whereas, in the Slovak population, a weak but significant deviation from HWE was found (FIS = 0.057, P=0.015). Further, after pooling the samples from both populations, a significant deviation from HWE was observed (FIS = 0.108, P<0.005). This suggests the existence of structure within the Czechoslovak otter population as a result of the Wahlund effect (Wahlund, 1928). Genetic differentiation of the populations was also indicated through the presence of a relatively high number of private alleles, and was confirmed by a highly significant FST value (FST = 0.154, P=0.0002). Using factorial correspondence analysis (FCA) plotting, samples from different localities were clustered into two distinct groups, clearly corresponding to separate Czech and Slovak populations. However, three individuals from the Czech population were plotted completely separately from both populations (Fig. 2). These three individuals were found to have originated from the area of the recent reintroduction project in the north-eastern part of the Czech Republic. The same results were obtained using Bayesian clustering implemented in programme STRUCTURE, i.e. the samples were divided into three groups: 129 individuals from the Czech Republic, 65 individuals from Slovakia, 3 individuals from the reintroduction area in the Czech Republic.
Demographic history
Using BOTTLENECK software, significant heterozygote excess was observed in both the Czech and Slovak populations when assuming infinite allele model (P<0.01), suggesting that otter populations in both countries declined recently. Further, both exponential and linear models using Beaumont’s (1999) procedure (MSVAR programme) also indicated a recent population decline in both populations. The results suggest that the decline in both populations started approximately 4-7 otter generations (i.e. 12-28 years) ago; however, the estimated parameters were slightly different between countries. Based on the exponential model which is more suitable for short timescale fluctuations, the effective population size (Ne) was assessed to be around 260 individuals before the decline started and 70 after the decline in the Czech population, and around 590 before and 150 after the decline in the Slovak population.Thus, both the pre- and the post-decline Ne were found to be lower in the Czech otter population, compared to that in the Slovak region, while the rate of decline was similar in both populations.
DISCUSSION AND CONCLUSIONS
Non-invasive genetic sampling
The success rate of non-invasive genotyping in our study was approximately 60%, which is a higher margin of the range observed in otter studies. Although Hansen and Jacobsen (1999) used PCR amplification of a part of the mitochondrial cytochrome b gene for species identification from less than one week old faeces with 100% success rate, amplification of microsatellite DNA is much more difficult process. Only very fresh spraints, or eventually old but frozen, have been found to yield amplifiable DNA for microsatellite genotyping, and the proportion of such samples ranged between 20-65% (Coxon et al., 1999; Dallas et al., 2003; Hung et at., 2004; Kalz et al., 2006; Arrendal et al., 2007; Ferrando et al., 2008, Koelewijn et al., 2010). Higher success rate (73%) was reported only by Janssens (2006) who selected preferentially jelly samples and viscous spraints for further analyses. This is consistent with the highest success rate of genotyping (82%) obtained for anal jelly samples in our study, as well as in some others (Coxon et al., 1999; Hájková et al., 2006; Lampa et al., 2008). The multiple-tubes approach (Taberlet et al., 1996) was essential to detect and correct genotyping errors and to produce reliable consensus genotypes.
At both study sites more males than females (1.2-1.4:1) were identified using NGS. Although this sex ratio did not differ significantly from 1:1, it can indicate higher ‘capture probability’ of males, resulting possibly from their higher mobility or higher marking activity. However, Koelewijn et al. (2010) reported the opposite in their non-invasive genetic monitoring study of the otter reintroduction in the Netherlands: females had significantly higher likelihood of being detected through spraints than males (91% vs. 65 %). It is also possible that such slightly biased sex ratio in our study can be a ‘random effect’, e.g. a consequence of the relatively small study site area (100 km2). An equal sex ratio was found in NGS study in otters on Kinmen (Hung et al., 2004), but a skewed sex ratio towards males (77% M:23% F) was reported from SW England (Dallas et al., 2003). Otter density was found to be 3.8-times higher in the fishpond region (Třeboňsko) compared to the oligotrophic mountainous area (Slovenský raj). Similarly, minimum home range sizes at Třeboňsko were much smaller (in hundreds meters) than in Slovenský raj (several kilometres). Non-invasive genetic sampling thus confirmed high density of otter population in Třeboňsko. However, also the density found in Slovenský raj, one otter per 3.8 km, is relatively high (especially for mountainous areas), although a similar density (one otter per 4-5 km) was reported from streams in Sweden (Erlinge, 1968) as well as from some Eastern European rivers (one otter per 1.7-5.8 km; Sidorovich et al., 1996; Sidorovich and Pikulik, 1998). It can be concluded, therefore, that food supply in streams of Slovenský raj is adequate. The high otter density in Třeboňsko is caused by large number of fishponds and channels providing abundant food supply. It makes this area a unique otter habitat in Europe. The non-invasive genetic method provided estimates of otter abundance in two different habitats and brought forward new information on their population structure and spatial distribution.
Genetic variability and population genetic structure
The genetic variability of microsatellite DNA in otter populations in the Czech and Slovak Republics was found to be similar, and within the range observed in other European otter populations (Dallas and Piertney, 1998; Dallas et al., 1999; Dallas et al., 2002; Pertoldi et al., 2001; Randi et al., 2003; Mucci et al., 2010). The relatively high FST value found in this study (0.15) is comparable with the FST value of 0.17 found on a much broader scale among several European otter populations (Randi et al., 2003), and is even slightly higher than the value found within large all-European study with 19 countries involved (FST = 0.13; Mucci et al., 2010). Such a level of genetic differentiation on a small geographic scale indicates a clear separation of the Czech and Slovak populations, confirmed also by FCA and Bayesian analysis. Perfect assignment of all individuals suggests that the populations may still be separated, i.e. there is no evidence for gene flow between the populations; however, no samples for analysis were obtained directly from the area of expected contact of populations.
Deviation from HWE in the Slovak population was only moderate and is not considered to be due to inbreeding. The Slovak population is connected with populations in Poland, Hungary and Ukraine, being part of the relatively continuous Central and Eastern European otter distribution range. A more probable explanation is the existence of further sub-structure within the Central and Eastern European population, which could possibly be detected if samples were also obtained from the surrounding countries.
The reason for the clear separation of the three otters from the two populations (Fig. 2) is due to the fact that at several loci, their genotypes carried unique alleles, present in neither the Czech nor Slovak populations. The carcasses of these three otters were found close to the area of reintroduction within which nine captive bred individuals, born at the Pavlov Otter Station (Czech Republic), were released. All nine individuals were offspring of one captive born female and two males from the South Bohemian population (A. Toman, pers. com.). Based on records from the Otter Studbook of the European Breeding Programme, the captive born female originated from the B-line captive otters from the Norfolk Wildlife Park (United Kingdom), with the origin of the paternal, and part of the maternal, lines in the wild otter population of England. The origin of the two ancestors in the maternal line, however, is not known (A. Melissen, pers. com.). Within a recent large-scale all-European study of genetic diversity and structure in otter populations, the same unique alleles were found in reintroduced otters in England and France, and in wild otters from Israel (E. Randi and N. Mucci, pers. com.). Successful breeding of the released individuals was confirmed through DNA analysis from the spraints of the parental otters living at the Pavlov Otter Station, as one of the carcasses had a genotype corresponding to the F2 generation. This finding confirms that successful breeding occurred within five years after the release of the first individuals. The reintroduction project, therefore, could theoretically be evaluated as successful (breeding success with wild individuals); however, in respect of preserving the original gene pool of the Czech otter population, individuals with an unknown origin should not have been released. Identification of those three otters with non-native origin also shows how well the genetic approach that we used works.
Demographic history
Both MSVAR and BOTTLENECK programmes indicated a very recent decline in both the Czech and Slovak otter populations. This decline is supported by the literature (e.g. Hell and Cimbal, 1978; Baruš and Zejda, 1981). According to the published data, the strongest decline probably occurred between the 1970s and the mid-1990s, very close to the estimate inferred by the MSVAR programme. The MSVAR programme indicated a similar recent demographic history for both populations, inferring a bottleneck-like decline to ca. 25% of the original Ne. The lower Ne inferred for the Czech population, as compared to the Slovak one, probably reflects the fact that the Czech population became isolated from any other otter population over the past century, while the Slovak population remained part of the more continuous Central and Eastern European otter population. The mean Ne estimates from the MSVAR programme seem to be fairly low, however, the ratio of Ne to census population size is often as low as 1/10 (Frankham, 1995).
Though this study did not reveal gene flow between the Czech and Slovak populations as yet, the populations are approaching each other and, in a few years, are expected to be fully re-connected and admixed. The Czech population in particular will benefit from this re-connection. A higher connectivity between the Czech and the Slovak populations will increase Ne, reduce the role of genetic drift, and increase genetic diversity. This will increase the otters’ evolutionary potential, i.e. the so-called ‘genetic rescue effect’ (Tallmon et al., 2004).
Despite the fact that otter populations are increasing in range at present, analysis of their demographic history indicates that their present effective population size is still very low, especially in the Czech Republic. The populations, therefore, remain highly vulnerable to any violent demographic change. While taking full account of the perceived problems of fish-farmers and anglers, it is highly desirable that otters remain a strongly protected species in both countries, and that the spreading and re-connection of presently isolated sub-populations is supported by habitat conservation. We suggest that, until the populations fully recover, and are once again able to sustain such pressures, emphasis is placed on economic tools and education in solving the present conflict between humans and otters.
This study was based on two types of non-invasively obtained material, i.e. tissue samples from carcasses and spraints of wild otters. It indicated that the use of ‘molecular scatology’ is highly effective for studies of genetic variability and population structure, particularly for elusive and protected species where the possibilities of obtaining samples are very restricted. Spraint samples significantly increased the amount of material available for this study and provided estimates of several population parameters which are very difficult to obtain in other way. However, as the genotyping of faecal DNA is connected with a high probability of genotyping errors, it remains necessary to follow the strict recommendations for non-invasive genetic sampling studies (e.g. Taberlet et al., 1999; Bonin et al., 2004; Smith et al., 2006).
ACKNOWLEDGMENTS - We thank Miroslav Lehocký, Vašek Bartuška and Jana Roche for help with collecting spraint samples in the field; and Zuzana Argalášová, Mária Boďová, Barbara Chovancová, Milan Ďurica, Zuzana Gálfyová, Ervin Hapl, Ladislav Hlôška, Viera Kacerová, Ján Kadlečík, Katarína Kisková, František Kunc, Anna Macková, Pavol Majko, Milan Olekšák, Marcela Roche, Olga Růžičková, Aleš Toman, Róbert Trnka, Andrea Ulehlová and Ivan Valach for providing, or making available, tissue samples from otter carcasses. We appreciate the advice and support concerning data analyses of Josef Bryja, Cino Pertoldi and Jan Zima. The study was supported by the Czech Science Foundation (grant no. 206/03/0757).
REFERENCES
Arrendal, J., Vila, C., Bjorklund, M. (2007). Reliability of noninvasive genetic census of otters compared to field censuses. Conserv. Genet. 8: 1097-1107.
Baruš, V., Zejda, J. (1981). The European otter (Lutra lutra) in the Czech Socialist Republic. Acta Sc. Nat. Brno 15 (12): 1-41.
Beaumont, M.A. (1999). Detecting population expansion and decline using microsatellites. Genetics 153: 2013-2029.
Belkhir, K., Borsa, P., Chikhi, L., Raufaste, N., Bonhomme, F. (1996-2004). GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions, CNRS UMR 5000, Université de Montpellier II, Montpellier (France).
Bonin, A., Bellemain, E., Bronken Eidesen, P., Pompanon, F., Brochmann, C., Taberlet, P. (2004). How to track and assess genotyping errors in population genetics studies. Mol. Ecol. 13: 3261-3273.
Broquet, T., Petit, E. (2004). Quantifying genotyping errors in noninvasive population genetics. Mol. Ecol. 13: 3601-3608.
Conroy, J.W.H., Chanin, P.R.F. (2002). The status of the Eurasian otter (Lutra lutra). IUCN Otter Spec. Group Bull. 19A: 24-48.
Cornuet, J.M., Luikart, G. (1996). Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 144: 2001-2014.
Coxon, K., Chanin, P., Dallas, J., Sykes, T. (1999). The use of DNA fingerprinting to study population dynamics of otters (Lutra lutra) in southern Britain: a feasibility study. R&D Technical Report W202, Environment Agency, Bristol, UK.
Dallas, J.F., Bacon, P.J., Carss D.N., Conroy J.W.H., Green, R., Jefferies D.J., Kruuk, H., Marshall, F., Piertney S.B., Racey, P.A. (1999). Genetic diversity in the Eurasian otter, Lutra lutra, in Scotland. Evidence from microsatellite polymorphism. Biol. J. Linn. Soc. 68: 73-86.
Dallas, J.F., Carss, D.N., Marshall, F., Koepfli, K.-P., Kruuk, H., Piertney, S.B., Bacon, P.J. (2000). Sex identification of the Eurasian otter Lutra lutra by PCR typing spraints. Conserv. Genet. 1: 181-183.
Dallas, J.F., Coxon, K.E., Sykes, T., Chanin, P.R.F., Marshall, F., Carss, D.N., Bacon, P.J., Piertney, S.B., Racey, P.A. (2003). Similar estimates of population genetic composition and sex ratio derived from carcasses and faeces of Eurasian otter Lutra lutra. Mol. Ecol. 12: 275-282.
Dallas, J.F., Marshall, F., Piertney, S.B., Bacon, P.J., Racey, P.A. (2002). Spatially restricted gene flow and reduced microsatellite polymorphism in the Eurasian otter Lutra lutra in Britain. Conserv. Genet. 3: 15-29.
Dallas, J.F., Piertney, S.B. (1998). Microsatellite primers for the Eurasian otter. Mol. Ecol. 7: 1248-1251.
Erlinge, S. (1968). Territoriality of the otter Lutra lutra L. Oikos, 19: 81-98.
Ferrando, A., Lecis, R., Domingo-Roura, X., Ponsa, M. (2008). Genetic diversity and individual identification of reintroduced otters (Lutra lutra) in north-eastern Spain by DNA genotyping of spraints. Conserv. Genet. 9: 129-139.
Frankham, R. (1995). Effective population size / adult population size ratios in wildlife: a review. Genet. Research 66: 95-107.
Frankham, R., Ballou, J. D., Briscoe, D. A. (2002). Introduction to Conservation Genetics. Cambridge University Press, Cambridge.
Goudet, J. (2001). FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.9.3). Updated from Goudet, 1995.
Hájková, P., Pertoldi, C., Zemanová, B., Roche, K., Hájek, B., Bryja, J., Zima, J. (2007). Genetic structure and evidence for recent population decline in Eurasian otter populations in the Czech and Slovak Republics: implications for conservation. J. Zool., Lond. 272: 1-9.
Hájková, P., Zemanová, B., Bryja, J., Hájek, B., Roche, K., Tkadlec, E., Zima, J. (2006). Factors affecting success of PCR amplification of microsatellite loci from otter faeces. Mol. Ecol. Notes 6: 559-562.
Hájková, P., Zemanová, B., Roche, K., Hájek, B. (2009). An evaluation of field and noninvasive genetic methods for estimating Eurasian otter population size. Conserv. Genet. 10: 1667-1681.
Hansen, M.M., Jacobsen, L. (1999). Identification of mustelid species: otter (Lutra lutra), American mink (Mustela vison) and polecat (Mustela putorius), by analysis of DNA from faecal samples. J. Zool., Lond. 247: 177-181.
Hell, P., Cimbal D. (1978). Rozšírenie a početnosť vydry riečnej (Lutra lutra L.) na Slovensku. [Distribution and abundance of the otter (Lutra lutra L.) in Slovakia]. Folia Venatoria 8: 223-236. [in Slovak with English summary]
Hung, C.-M., Li, S.-H., Lee, L.-L. (2004). Faecal DNA typing to determine the abundance and spatial organisation of otters (Lutra lutra) along two stream systems in Kinmen. Anim. Conserv. 7: 301-311.
Janssens, X. (2006). Monitoring and predicting elusive species colonisation. Application to the otter in the Cévennes National Park (France). PhD Thesis. Université catholique de Louvain, Louvain-la-Neuve, Belgium.
Kalz, B., Jewgenow, K., Fickel, J. (2006). Structure of an otter (Lutra lutra) population in Germany - results of DNA and hormone analyses from faecal samples. Mammal. Biol. 71: 321-335.
Koelewijn, H.P., Pérez-Haro, M., Jansman, H.A.H., Boerwinkel, M.C., Bovenschen, J., Lammertsma, D.R., Niewold, F.J.J., Kuiters, A.T. (2010). The reintroduction of the Eurasian otter (Lutra lutra) into the Netherlands: hidden life revealed by noninvasive genetic monitoring. Conserv. Genet. 11: 601-614.
Kohn, M.H., Wayne, R.K. (1997). Facts from feces revisited. Trends Ecol. Evol. 12: 223-227.
Lampa, S., Gruber, B., Henle, K., Hoehn, M. (2008). An optimisation approach to increase DNA amplification success of otter faeces. Conserv. Genet. 9: 201-210.
Macdonald, S.M., Mason, C.F. (1994). Status and conservation needs of the otter (Lutra lutra) in the western Palearctic. Nature and Environment. No. 67. Council of Europe Press, Strasbourg.
Mason, C.F., Macdonald, S.M. (2004). Growth in otter (Lutra lutra) populations in the UK as shown by long-term monitoring. Ambio 33: 148-152.
Mills, L.S. (2007). Conservation of Wildlife Populations. Demography, Genetics and Management. Blackwell Publishing, Malden, Oxford, Carlton.
Mucci, N., Arrendal, J., Ansorge, H., Bailey, M., Bodner, M., Delibes, M., Ferrando, A., Fournier, P., Fournier, C., Godoy, J.A., Hajkova, P., Hauer, S., Heggberget, T.M., Heidecke, D., Kirjavainen, H., Krueger, H.-H., Kvaloy, K., Lafontaine, L., Lanszki, J., Lemarchand, C., Liukko, U.-M., Loeschcke, V., Ludwig, G., Madsen, A.B., Mercier, L., Ozolins, J., Paunovic, M., Pertoldi, C., Piriz, A., Prigioni, C., Santos-Reis, M., Luis, T.S., Stjernberg, T., Schmid, H., Suchentrunk, F., Teubner, J., Tornberg, R., Zinke, O., Randi, E. (2010). Genetic diversity and landscape genetic structure of otter (Lutra lutra) populations in Europe. Conserv. Genet. 11: 583-599.
Paetkau, C., Strobeck, C. (1994). Microsatellite analysis of genetic variation in black bear populations. Mol. Ecol. 3: 489-495.
Pertoldi, C., Hansen, M.M., Loeschcke, V., Madsen, A.B., Jacobsen, L., Baagoe, H. (2001). Genetic consequences of population decline in the European otter (Lutra lutra): an assessment of microsatellite DNA variation in Danish otters from 1883 to 1993. Proc. R. Soc. Lond. B, 268: 1775-1781.
Piry, S., Alapetite, A., Cornuet, J.M., Paetkau, D., Baudouin, L., Estoup, A. (2004). GeneClass2: a software for genetic assignment and first generation migrants detection. J. Hered. 95: 536-539.
Pritchard, J., Stephens, M., Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155: 945-959.
Randi, E., Davoli, F., Pierpaoli, M., Pertoldi, C., Madsen, A.B., Loeschcke, V. (2003). Genetic structure in otter (Lutra lutra) populations in Europe: implications for conservation. Anim. Conserv. 6: 1-10.
Reuther, C., Krekemeyer, A. (2004). Digital map of the distribution of the Eurasian otter (Lutra lutra) in Europe from 1990 to 2003, based on the UTM 10km-grid and the ’Information System Otter Signs’ (ISOS). In On the way towards an Otter Habitat Network Europe (OHNE). C. Reuther, A. Krekemeyer (eds.), Schriftenreihe Habitat: Arbeitsberichte der AKTION FISCHOTTERSCHUTZ e. V. 15:, 308 pp.
Roche, K., Toman, A., Šusta, F. (2004). National otter (Lutra lutra) survey of the Czech Republic 1997-2002(3). In Scientific report of the Czech Otter Project 1998-2004. K. Roche (ed.), Czech Otter Foundation Fund, Trebon, pp. 14-29.
Sidorovich, V.E., Jedrzejewska, B., Jedrzejewski, W. (1996). Winter distribution and abundance of mustelids and beavers in the river valleys of Bialowieza Primeval Forest. Acta Theriol. 41: 155-170.
Sidorovich, V.E., Pikulik, M.M. (1998). Factors allowing high density of otters in Eastern Europe. In Proceedings of the 7th International Otter Colloquium: Otter Conservation - An Example for a Sustainable Use of Wetlands. R. Dulfer, J. Conroy, J. Nel, A. Gutleb (eds.), IUCN Otter Spec. Group Bull. 19A: 326-333.
Smith, D.A., Ralls, K., Hurt, A., Adams, B., Parker, M., Maldonado, J.E. (2006). Assessing reliability of microsatellite genotypes from wild kit fox faecal samples usány genetic and GIS analyses. Mol. Ecol. 15: 387-406.
Storz, J.F, Beaumont, M.A. (2002). Testing for genetic evidence of population expansion and contraction: an empirical analysis of microsatellite DNA variation using a hierarchical Bayesian model. Evolution 56: 154-166.
Taberlet, P., Griffin, S., Goossens, B., Questiau, S., Manceau, V., Escaravage, N., Waits, L.P., Bouvet, J. (1996). Reliable genotyping of samples with very low DNA quantities using PCR. Nucleic Acids Research 24: 3189-3194.
Taberlet, P., Luikart, G. (1999). Non-invasive genetic sampling and individual identification. Biol. J. Linn. Soc. 68: 41-55.
Taberlet, P., Waits, L.P., Luikart, G. (1999). Noninvasive genetic sampling: look before you leap. Trends Ecol. Evol. 14: 323-327.
Tallmon, D., Luikart, G., Waples, R.S. (2004). The alluring simplicity and complex reality of genetic rescue. Trends Ecol. Evol. 19: 489-496.
Urban, P., Kadlečík, J. (2001). Program záchrany chráneného ohrozeného druhu vydra riečna Lutra lutra (Linnaeus, 1758). [Action plan of protected endangered species, the Eurasian otter Lutra lutra (Linnaeus, 1758)]. Štátna ochrana prírody SR, Banská Bystrica, 20 pp. [in Slovak]
Valière, N., Berthier, P., Mouchiroud, D., Pontier, D. (2002). GEMINI: software for testing the effects of genotyping errors and multitubes approach for individual identification. Mol. Ecol. Notes 2: 83-86.
Wahlund, S. (1928). Composition of populations from the perspective of the theory of heredity. Hereditas 11: 65-105.
Waits, L.P., Luikart, G., Taberlet, P. (2001). Estimating the probability of identity among genotypes in natural populations: cautions and guidelines. Mol. Ecol. 10: 249-256.
Résumé :Genetique de la Conservation et Echantillonnage Genetique de la Loutre (Lutra lutra) en Republiques Tcheque et Slovaque
De nombreux paramètres importants pour la gestion et la réussite de conservation des populations de Loutre (Lutra lutra) ne sont pas encore parfaitement compris. Les loutres sont des animaux rares et insaisissables, et il est très difficile d'étudier leur abondance et la structure des populations. Toutefois, au cours des dernières années, des méthodes génétiques ont permis une nouvelle approche du problème. L’obtention d’échantillons de façon non-invasive telles que les matières fécales permet de pas ne déranger ou même observer les animaux. Nous avons utilisé une méthode génétique non-invasive pour évaluer la taille de la population, le sex-ratio et la répartition spatiale des loutres dans deux habitats différents. De plus, nous avons étudié la structure génétique des loutres dans les républiques tchèque et slovaque en utilisant l'ADN extrait à la fois de tissus et d’épreintes. Un total de 448 épreintes et de 134 tissus ont été analysés en utilisant 10 loci microsatellitaires et un marqueur SRY. Sur le premier site d’étude, une zone montagneuse du parc national de Slovenský raj, 13 individus (7 mâles et 6 femelles) ont été identifiés, tandis que sur le deuxième site, une zone d’étangs de production dans la réserve de biosphère de Třeboň, ce sont 50 individus (29 mâles et 21 femelles) qui ont été recensés. Les deux sites faisant la même taille, environ 100 km2. Lorsque l'on étudie la structure génétique des populations de loutres dans les deux républiques, un niveau relativement élevé de différenciation se distingue entre les populations (FST = 0,154), ceci étant soutenu par une scission parfaite des clusters bayésiens. Les deux programmes MSVAR et BOTTLENECK qui déduisent l'histoire démographique des populations, indiquent l'apparition récente de goulets d'étranglement démographiques dans chacune des populations. Malgré la recolonisation récente, les populations de loutres sont encore vulnérables dans les deux pays.
Revenez au dessus
Resumen: Conservación Genética y Muestreo Genético No Invasivo en Nutria Euroasiática (Lutra lutra) en las Republicas Checa y Eslovaca
Varios parámetros básicos e importantes de la nutria euroasiática (Lutra lutra) aún no son totalmente entendidos para un exitoso manejo y conservación de esta especie. Las nutrias son animales raros y elusivos, y es muy difícil estudiar su abundancia y estructura poblacional. Sin embargo, durante los últimos años los métodos genéticos están proveyendo una nueva aproximación a este problema. Mediante la colección de muestras no invasivas, tales como heces, estas son analizadas y así el disturbio u observación de los animales no es requerido. Nosotros utilizamos un método no invasivo para evaluar la edad de la población, razón de sexo y distribución espacial de nutrias en 2 ambientes distintos. Además, nosotros estudiamos la estructura genética de nutrias en las Repúblicas Checa y de Eslovaca usando ADN extraído desde muestras de tejido y heces. Un total de 448 heces y 134 tejidos fueron analizados usando 10 microstalites loci y un marcador SRY. En el primer área de estudio, en la zona montañosa del Parque Nacional Slovenský raj, 13 individuos(7 machos y 6 hembras) fueron identificados, mientras que en el segundo sitio, un estanque de peces de la Reserva de la Biosfera de Třeboňsko, con el mismo tamaño de área (100 km2) había 50 individuos (29 machos y 21 hembras). Cuando se analizó la estructura genética de las poblaciones de las Republicas Checa y Eslovaca, se encontró una diferenciación genética (FST = 0.154), apoyada por una asignacion perfecta en el agrupamiento Bayesiano. Los programas MSVAR y BOTTLENECK, los cuales infieren historia demográfica, indicaron la ocurrencia de un cuello de botella poblacional reciente en ambas poblaciones. A pesar de la reciente recuperación, las poblaciones de nutrias en ambos países estas todavía vulnerables.
Vuelva a la tapa