IUCN/SSC Otter Specialist Group Bulletin

|
©IUCN/SCC Otter Specialist Group Volume 28A Proceedings Xth International Otter Colloquium, Hwacheon, South Korea Citation: Franco, M. and Soto-Gamboa, M. (2011) Prey Selection And Trophic Position Of Southern River Otter (Lontra provocax) In The Wetlands of the “Carlos Adwandter Nature Sanctuary”, Southern Chile. Proceedings of Xth International Otter Colloquium, IUCN Otter Spec. Group Bull. 28A: 86 - 96 Prey Selection And Trophic Position Of Southern River Otter (Lontra provocax) In The Wetlands of the “Carlos Adwandter Nature Sanctuary”, Southern Chile Marcela Franco and Mauricio Soto-Gamboa
1Instituto de Ciencias de la Tierra y Evolución, Facultad de Ciencias, Universidad Austral de Chile. Campus Isla Teja, Casilla 567, Valdivia, Chile |

 |
| (Received 23rd March 2011, accepted 13th August 2011) |
|
Abstract: Otters are considered as top predators and participate in ecosystem services and affect community structure by the modulation of prey densities. Nevertheless, diet of the southern river otter (Lontra provocax) has been described only based on the frequency of occurrence of prey in feces. Actually, the relationship between trophic availability and prey selection is unknown. In this study we evaluated the relationship between prey selection and availability in different environments. We hypothesized that this species is an opportunistic predator, and that their diet could be determined by prey abundance. We studied fecal remains in two different environments (marsh–river) in the “Carlos Andwanter Nature Sanctuary” in southern Chile. The prey found in the scats were identified through reference collections. We estimated the frequency of occurrence, relative availability of prey item, and the Selection Index. In addition, we quantified prey availability using electrofishing. Our results suggest that the trophic niche for L. provocax is restricted to a few prey items, with crayfish (Samastacus spinifrons) as the most important diet component. However, L. provocax consume specific crayfish size independently of their availability in the environment. Our results suggest that L. provocax is an opportunistic predator, but shows a clear selection of medium-sized crayfish. |
| Keywords: top predator, wetland, southern river otter, Lontra provocax, trophic selection. |
| Française | Español |
INTRODUCTION
Otters are carnivore mustelids recognized as top predators of aquatic ecosystems, capable of shaping the community structure through their trophic interactions (Anoop and Hussain, 2005). Current knowledge of river otter trophic ecology is based on long-term studies of the Eurasian Lutra lutra (Kruuk and Moorhouse, 1990,1991; Kruuk et al., 1987; Heggberget, 1993), and the North American Lontra canadensis (Melquist and Hornocker, 1983, Blundell et al., 2002) and more recent studies on Lontra longicaudis (Pardini, 1998), Lontra perspicillata (Anoop and Hussain, 2005), Aonyx capensis (Somers, 2000), and Lontra provocax (Medina, 1998). The diet of Latin American otters is only known from faecal analysis, except for Lontra longicaudis for which Pardini (1998) integrated information on prey availability. Some authors discussed the importance of estimating prey availability to determine the actual feeding preferences of otters, because it is not possible to do this with fecal analysis alone (Pardini, 1998; Anoop and Hussain, 2005). Moreover, the availability of resources and the evaluation of predator-prey relationships is a prerequisite for understanding factors that limit populations, for assessing competition, and for designing conservation management strategies, as the otters face changes in prey availability in its habitat (Kruuk, 2006).
Based on their feeding ecology, otters are regarded as opportunistic predators, due to their high dietary variability between habitats (Chanin, 1985), and their diet in relation to prey availability (Pardini, 1998). However, there is growing evidence showing that otters have preferences in prey type and size, vulnerability, abundance and habitat (bottom-living species) (Reid et al., 1994, Carter and Rosas, 1997; Medina, 1998; Pardini, 1998; Somers, 2000; Quadros and Monteiro, 2001; Anoop and Hussein, 2005; Kruuk, 2006). A recent study on Enhydra lutris demonstrates that this otter has higher foraging costs with prolonged periods of rest on the water surface (Finerty et al., 2009). Therefore, trade-off between hunting, handling techniques and food processing can explain the foraging preferences of otters (Yeates et al., 2007).
Some authors proposed that otters are top predators able to modulate community structure and prey density of aquatic ecosystems (Anoop and Hussein, 2005). However, this has been tested only for Enhydra lutris, the sea otter that regulate sea urchin populations and algal communities in Alaskan rocky habitats and in Southern California (Estes and Duggins, 1995). Initially, studies related to feeding ecology have been conducted using stomach contents and faecal or food remains. However, this information is descriptive and does not incorporate spatial and temporal factors implicated in the trophic interactions between community components (Angerbjörn et al., 1994).
The Southern river otter (Lontra provocax) of southern Chile and Argentina is classified as endangered on the IUCN Red List of Threatened Species (IUCN 2002). Historically, the distribution was from 34° S to 53°S in Chile, and from the Andes to the Pacific (Osgood, 1943; Medina, 1996; Medina et al., 2003). Today, the populations in freshwater habitats are restricted to a few isolated areas, from 39°S to 44° S (Medina, 1996). The major factors leading to population decline is habitat degradation, rivers canalization and drainage, decrease of preys, and furtive hunting; all of which has drastically affected the distribution patterns of this species(Medina 1996, Medina et al., 2003).
Knowledge about the feeding ecology of Southern river otters is fragmented and comes from descriptive studies of faecal analysis. Therefore, the relationship between prey consumed and their availability in the environment, as well as the foraging strategies of this endangered otter are unknown. We hypothesized that because the Southern river otter has been described as an opportunistic predator, its diet will reflect the availability of the different prey species in the environment. The aims of this study are: 1) determine the relationship between diet and prey availability; 2) establish whether trophic preferences or selection of preys according to size and species exists; and 3) determine the foraging strategies used by Southern river otters.
METHODS
Study Area
Our study area was located in the “Carlos Andwanter Nature Sanctuary”, located between the fork of the Cruces and Cau-Cau rivers near Valdivia city (39°49'S, 73°15'W); this area includes the wetland “Cruces River” and its affluent (Fig. 1). The superficial area cover is approximately 4877 ha (Schlatter et al., 1991). The Cruces River is the principal channel of the wetland system, which includes a marsh area “hualve”, which is a shallow flooding system with muddy substratum and an emergent vegetative association of Myrtaceae that generate a heterogeneous habitat subjacent to the river. In the principal channel, the waters are calm and depth ranges from 4 to 8 m, with a silt and sandy substratum (Steubing et al., 1980; Ramírez et al., 1991). The sanctuary was designated as a RAMSAR (Convention on Wetlands of International Importance) site on 27 July 1981 (Corti and Schlatter, 2002).
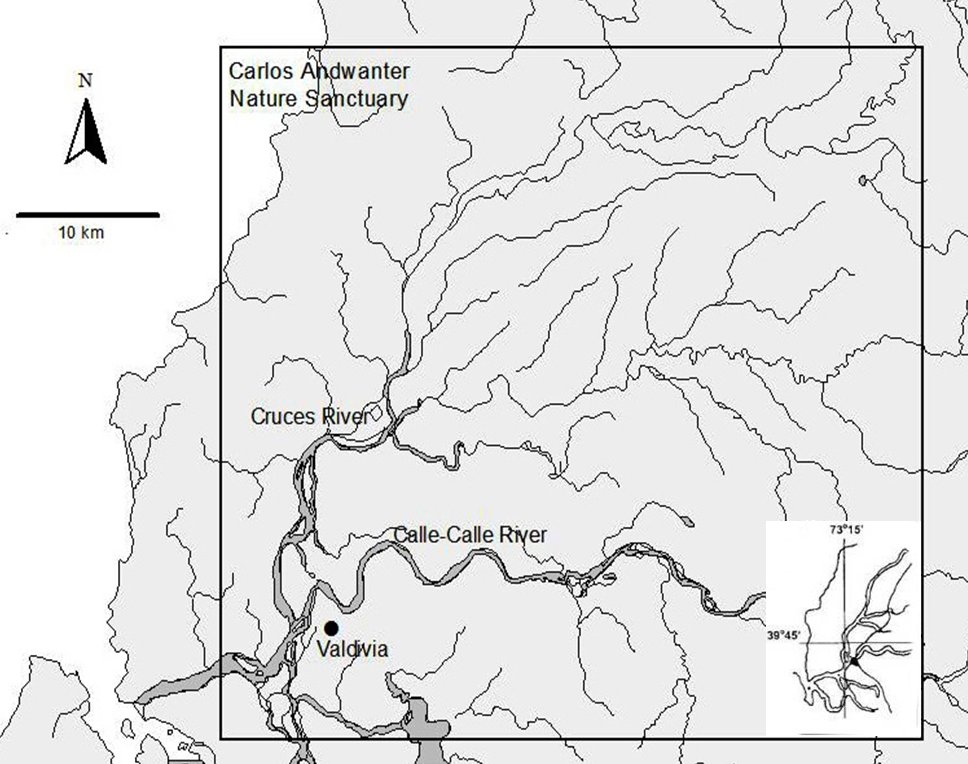 |
| Figure 1. Location of the wetlands of the “Carlos Andwanter Nature Sanctuary” (click for larger version) |
Spraint Collection
Spraints were collected in two habitats in the sanctuary (marsh and river), where burrows and latrine sites were identified. These sites were regularly surveyed along 30 km of river banks and the marsh during summer (December - March) 2006. The spraints collected were stored individually for later analysis in the laboratory.
Faecal Analysis and Diet Quantification
Spraints were washed under tap water in a sieve of mesh size 0.5 mm. They were dried in the shade, and the undigested parts were separated and identified using a reference collection from the Instituto de Zoología (Universidad Austral de Chile). The vertebra and venation patterns of the scales were used to identify fish species. The sharp lateral spines of the catfish (Dyplomystes spp.) made its identification easy, while crustacean species were identified from the colour and diagnostic shapes of exoskeleton remains. Dietary composition was reported as frequency of occurrence (i.e., number of spraints in which a species occurred divided by the total number of spraints collected) and relative frequency (i.e., number of spraints in which a species occurred divided by the total occurrence of all the species tested) of the prey in the spraints (Erlinge, 1968). Differences between habitats in the diet were evaluated by Kruskal-Wallis analysis (Zar, 1999).
Prey Availability
Crustacean and fish density (ind. m-2) was evaluated using electrofishing. We selected three sampling sites in each habitat (river and marsh); the distance of transects was determined accord to the physical features of sites (e.g. width and depth), but in general ≤25 meters and took less than ten minutes to complete. Logistical problems and difficulty of access to sampling sites did not allow us to obtain more sampling sites for electrofishing in each habitat. Differences in prey availability between habitats were evaluated by Kruskall-Wallis analysis (Zar, 1999).
Estimation of Prey Size from Spraint Remains
Crustaceans collected during sampling were used. Prey size found in the diet was estimated using linear regression between morphometric variables, body length vs. telson length and body length vs. uropods length. We measured the same variables from remains of S. spinifrons found in the spraints. After this, we replaced each value (telson and uropod length) in the linear regression equation obtained by that of the shrimp caught in the field.
Preference and Prey Selection.
We used the index of Manly (1995) to determine the preference between the most abundant prey species (S. spinifrons, Aegla spp. and fishes).
| Manly (1995) Index: | 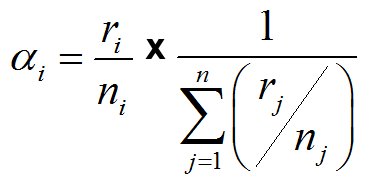 |
Where ri = % prey in the diet, and ni = % in the environment, and rj =% of the next prey in the diet, and nj =% of the that prey in the environment. After, we estimated the trophic selection index Chesson (1983) to establish the selectivity of each prey in relation to its availability in the environment.
Chesson (1983) trophic selection index: |
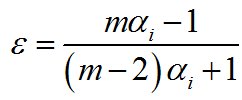 |
Where αi= Manly Index of species i, m = number of prey evaluated. We used the values of frequency of occurrence in the diet and in the environment.
RESULTS
Diet of Lontra provocax
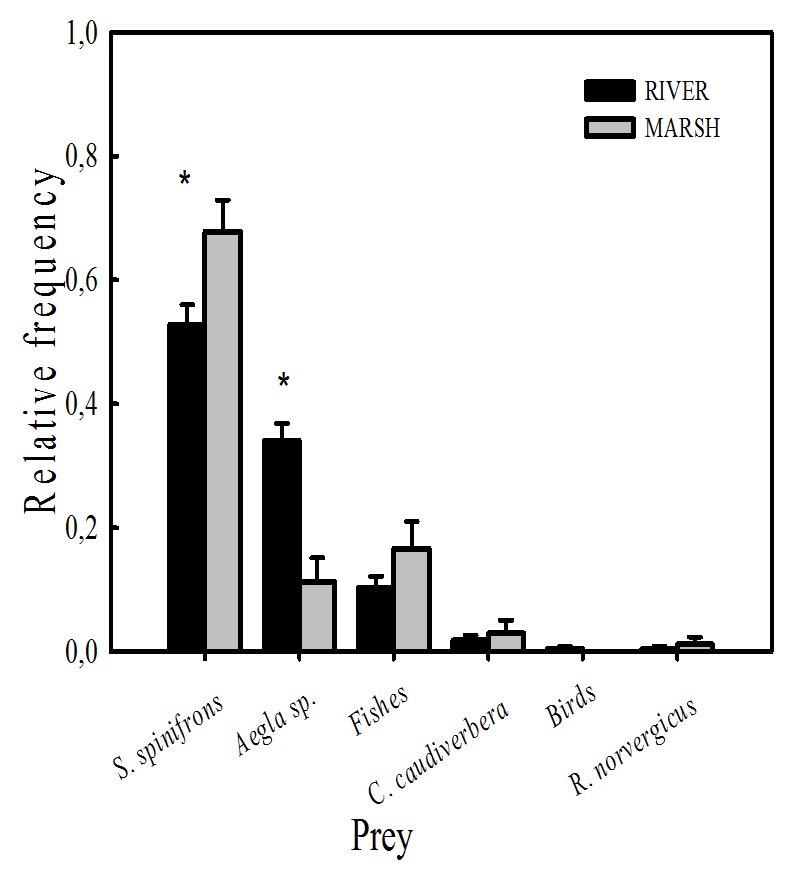
We found that crustaceans constitute the major component of the diet of the southern river otter, mostly crayfish Samastacus spinifrons (93.58% in river and 100% in marsh), as well as crabs Aegla spp. (73.07% in river and 21.42% in marsh). Aegla spp. was more frequent in the river than in the marsh (Table 1). We did not found significant differences in the frequency of occurrence of fishes between habitats.
We found significant differences between habitats in the frequency of crustaceans consumed, S. spinifrons (Kruskall-Wallis H (1,106) = 5.558, P<0.02) and Aegla spp. (Kruskall-Wallis H (1,106) = 15.793, P<0.001) (Fig. 2).
Availability of Crustaceans Consumed by L. Provocax in the Environment
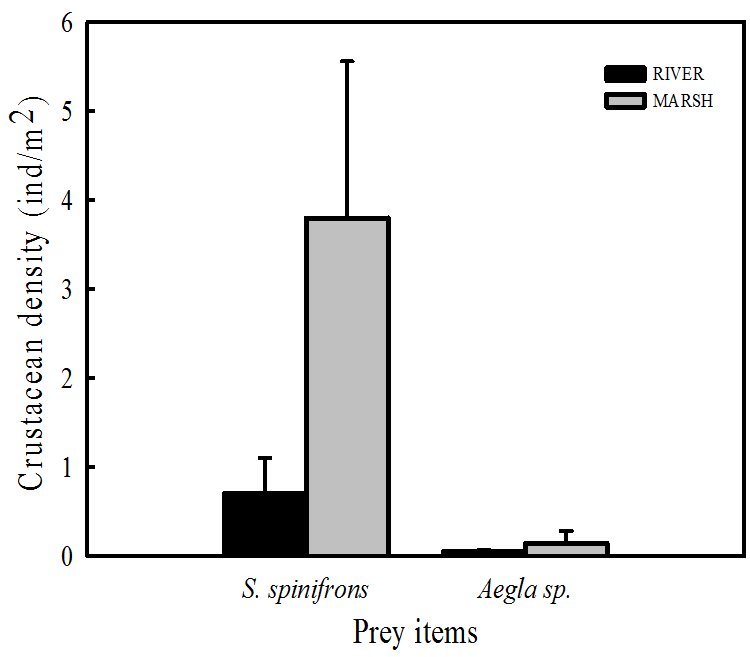
Significant differences between habitats were found for S. spinifrons (Kruskal-Wallis H (1,7) = 4.125, P<0.05) but not for Aegla spp. (Kruskal-Wallis H (1,7) = 0.134, P>0.05) (Fig. 3).
Estimation of Crayfish Size from Remains in the Spraints
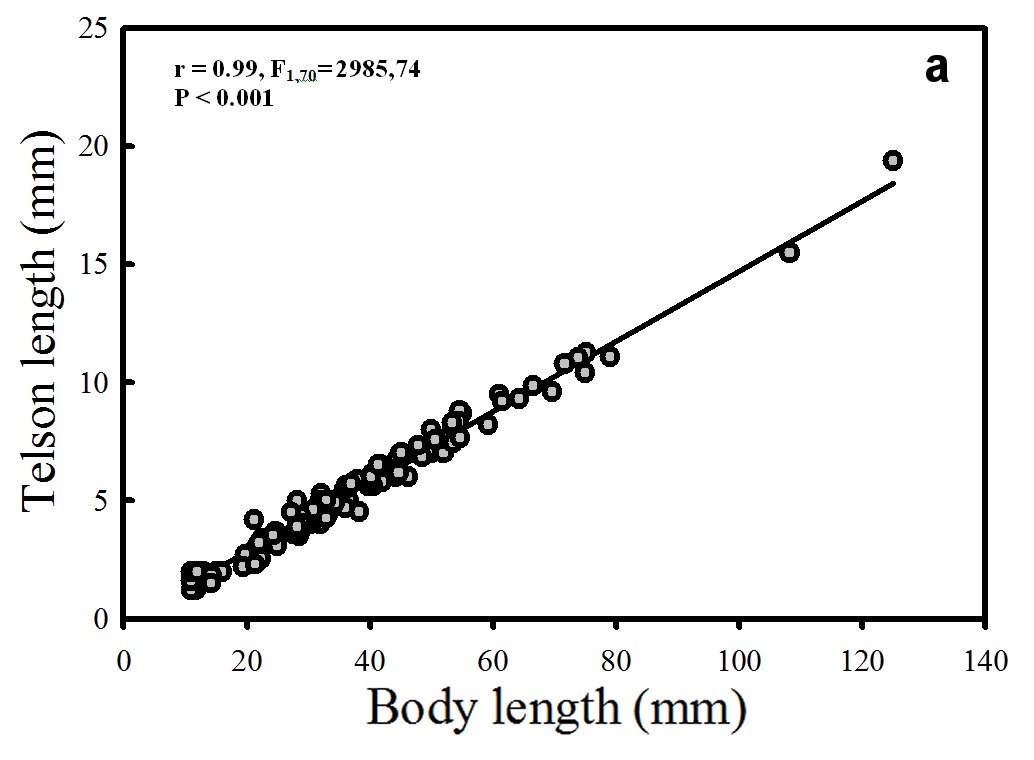
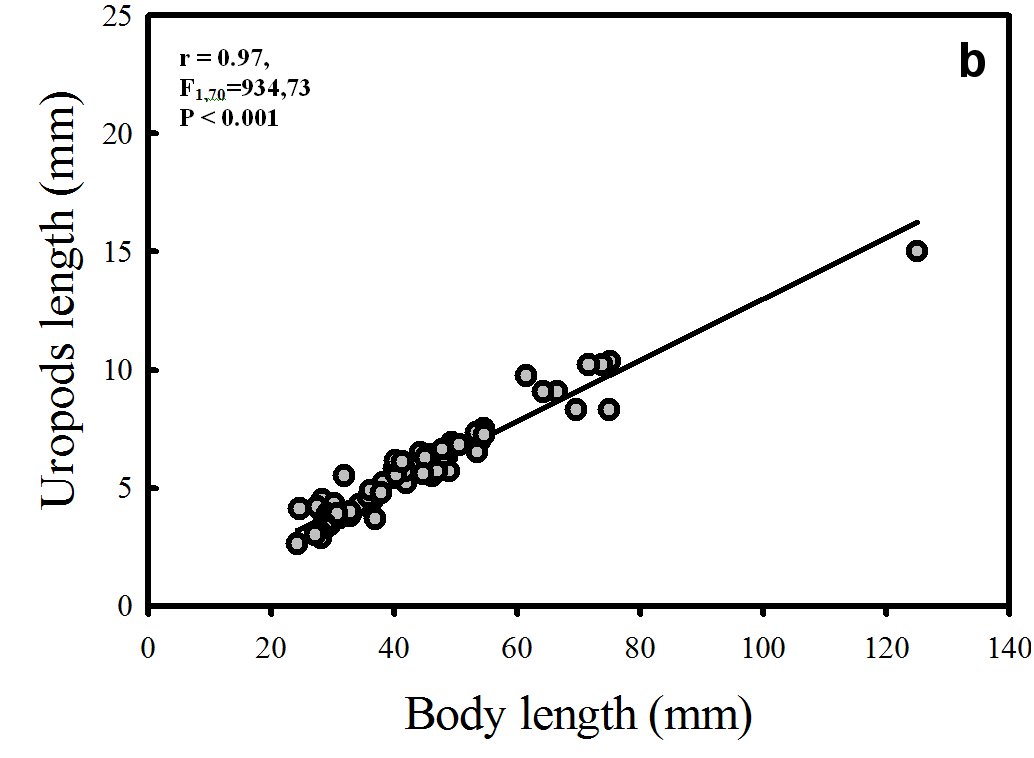
A positive correlation (significant linear regression) between the independent variable body length (mm) and the dependent variables (x), telson length (Fig. 4a) and uropod length (mm) was found (Fig. 4b).
Size of Samastacus spinifrons (mm) in the Diet and the Environment
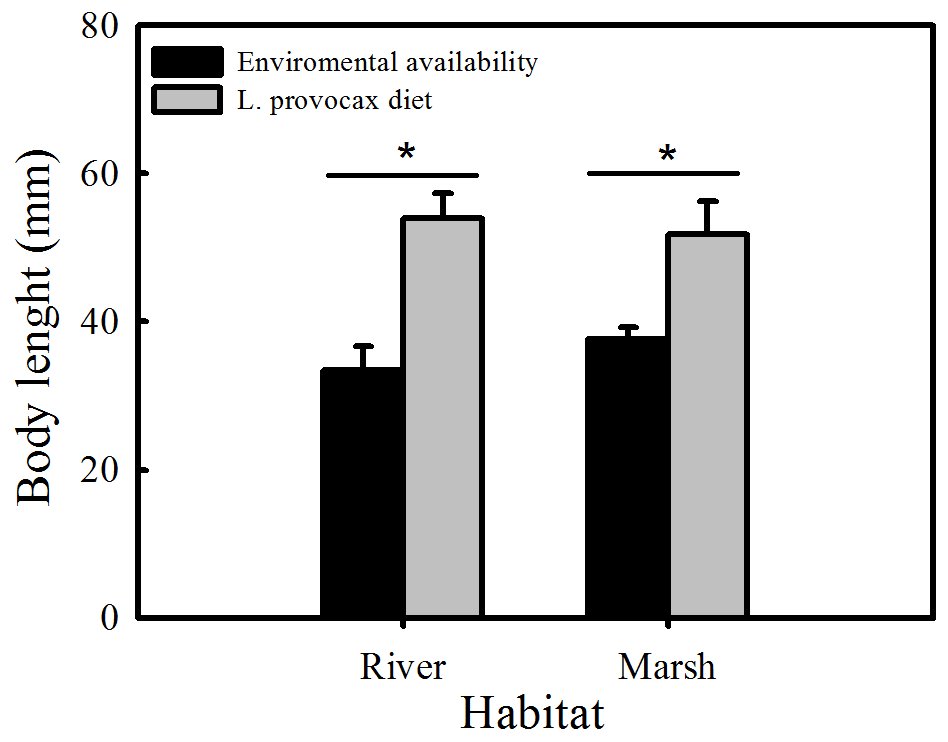
No significant differences between habitats were found for crayfish size (body length) either in the diet or in the environment (Kruskal-Wallis H (1,48) = 0.0922, P>0.05) (Fig. 5), but significant differences between availability and consumption by L. provocax in both environments were detected (ANOVA, F1,34 = 4.1910, P<0.05).
Trophic Selectivity Index in the River
Our results suggest that the Southern river otter prefer crustaceans over fishes by a large margin. Among crustaceans, they seem to select S. spinifrons, which was also more abundant in the environment (Fig. 3). Lontra provocax select prey from highest values to lowest isotopic values, S. spinifrons (ε=0.98) and Aegla sp. (ε=0.85) while fish (ε=-0.89) were little selected. Otters were feeding mainly on slow-moving and bottom-dwelling fishes such as cyprinids and catfishes, and to a lesser degree on pelagic fish such as salmonids.
DISCUSSION
As has been shown for other otter species (Somers, 2000), and as is evident from other studies (Chéhebar et al., 1986; Medina, 1997; 1998), crustaceans are important prey for Lontra provocax in freshwater habitats in South Chile. Fish, mainly cyprinids, catfish, and to a lesser extent some salmonids, are also frequent in the diet. Frogs, birds, and rodents are found at very low frequencies. Crustaceans and fishes have also been reported as the two most important food items for this species in lentic habitats (Medina, 1997, 1998). In the present study, we find that Southern river ottershow dietary preferences independent of habitat, and they seem to select both crustaceans and fish in relation to their position in the water body (depth), prey microhabitat and their low escape ability.
Crustaceans have been described as having low energetic content compared to fish (Kruuk, 1995, 2006). Our results suggest that the Southern river otter is heavily dependent on crustaceans and selects S. spinifrons; which like Aegla spp., is benthic (Jara, 1994). This preference may be related to greater abundance of crayfish with respect to crabs and fish. Among fish, otters also prefer the bottom-dwelling cyprinids and catfish over those that are active swimmers in the water column. Once again, otter preferences may be related to the low escape ability of prey in its dwelling place. Erlinge (1968), studying prey selection by L. lutra in captivity, concluded that otters catch fish in inverse proportion to their ability to escape. The few diet studies on L.provocax do not analyse prey availability. However, the high predation rates on crayfish S. spinifrons, crab Aegla rostrata and salmonids at “Todos los Santos” and “Panguipulli” lakes (Medina, 1998), indicate that otters are able to capture these mobile fishes often, at least in lentic habitat.
We found that prey size is also a factor influencing prey selection for otters. Some studies that have analyzed prey availability show that otters select on both prey size and species (Van der Zee, 1981; Wise et al.,1981; Kruuk and Moorhouse, 1990; Pardini, 1998). This can explain the mean size of crayfish eaten for the otters, considering that these reach a size of up to 147 mm in body length (Rudolph, 2002). It is probable that escape ability, abundance, and size are not the only factors influencing prey selection of Southern river otter. Factors such as prey microhabitat, conspicuousness and palatability also seem to influence prey selection in Lutra lutra (Kruuk and Moorhouse, 1990; Wise et al.,1981; Kruuk, 2006).
Structural heterogeneity of river and marsh “hualve” provide a suitable refuge for crustaceans, especially for crayfish S. spinifrons, which live in backwaters between riparian vegetation, where there are shallow caves and refuges under submerged trunks, roots, and rocks (Jara, 1994; Rudolph 2002). Thus, microhabitat factors may influence the efficiency of otters to catch their prey (Kruuk, 2006). Factors such as water depth, riparian vegetation and root availability may be most important (Mason and Macdonald 1986; Chanin, 1985; Kruuk and Moorhouse, 1990; Kruuk, 1995; Pardini 1998; Medina, 1998; Medina et al.,2003). This agrees with our results, as the marsh presented a great availability of S. spinifrons, which is coherent with the high frequency of this prey in the Southern river otter diet.
The trophic niche of L. provocax is restricted to a few food items, where crayfish (S. spinifrons) is the most important component in the diet. Our results suggest that the Southern river otter is an opportunistic predator, eating prey in relation to their availability in the environment. However, it is selective opportunist and selects its prey mainly in relation to prey size, as well as slow moving and bottom-living prey, which is independent of habitat in the “Carlos Adwandter Nature Sanctuary”. Therefore, we suggest that the foraging strategy of L. provocax involves optimization of net rate of food intake as a function of catch efficiency. Because crayfish has low energy content, it will be selected for high abundances and low catch and handling costs. We thus accept the hypothesis that the L. provocax dietwill reflect the availability of the prey species in the environment. In summary, we suggest that the southern river otter is a selective opportunist predator to consume benthic detritivorous preys, and these preferences are associated to availability more than energetic quality of prey.
ACKNOWLEDGMENTS - We thank CONAF (Corporación Nacional Forestal) and the gamekeeper Luis Miranda and Luis Tón for their assistance in the field, collecting scats and helping with orientation in the Sanctuary. We are also grateful to CODEFF (Comité Nacional Pro Defensa de la Fauna y Flora), especially to Jose Luis Bartheld and Maximiliano Sepúlveda. Also, we thank Dr. Carlos Jara for assistance with crustacean identification. Marcela Franco would like to thank CONICYT doctoral fellowship (Comisión Nacional de Investigación Científica y Tecnológica), and Mauricio Soto-Gamboa to MECESUP AUG-011.
REFERENCES
Angerbjörn, A., Hersteinsson, P., Kerstin, L., Nelson, E. (1994). Dietary variation in arctic foxes (Alopex lagopus)- an analysis of stable carbon isotopes. Oecologia. 99: 226-232.
Anoop, K. R., Hussain, S. A. (2005). Food and feeding habits of smooth-coated otters (Lutra perspicillata) and their significance to the fish population of Kerala, India. Journal of Zoology. 266: 15-23.
Blundell, G. M., Ben-David, M., Bowyer, T. (2002). Sociality in river otters: cooperative foraging or reproductive strategies?. Behavioral Ecology. 13: 134-141.
Carter, S. K., Rosas, F. C. (1997). Biology and conservation of the giant otter Pteronura brasiliensis. Mammal Review. 27: 1–26.
Chanin, P. (1985). The natural history of otters. London: Croom Helm.
Chesson, J. (1983). The estimation and analysis of preference and its relationship to foraging models. Ecology. 64: 1297-1304.
Chéhebar, C.E., Gallur, A., Giannico, G., Gotelli, M., Yorio, P. (1986). A survey of the Southern river otter Lutra provocax in Lanin, Puelo and Los Alerces Nacional Parks, Argentina, and evaluation of its conservation status. Biological Conservation. 38: 293-304.
Corti, P., Schlatter, R. (2002). Feeding Ecology of the Black-necked Swan Cygnus melancoryphus in two Wetlands of Southern Chile. Studies on Neotropical Fauna and Environment. 37: 9-14.
Erlinge, S. (1968). Food studies on captive otters Lutra lutra L. Oikos. 19: 259-270.
Estes, J. A., Duggins, D. O. (1995). Sea otters and kelp forests in Alaska: Generality and variation in a community ecological paradigm. Ecological Monographs. 65: 75-100.
Finerty, S. E., Wolt, R. C., Davis, R. W. (2009). Summer activity pattern and field metabolic rate of adult male sea otters (Enhydra lutris) in a soft sediment habitat in Alaska. Journal of Experimental Marine Biology and Ecology. 377: 36-42.
Heggberget, T. M. (1993). Marine-feeding otters (Lutra lutra) in Norway: seasonal variation in prey and reproductive timing. Journal of Marine Biology and Assessment. 73: 297-312.
IUCN. (2002). Red List of Threatened Species. IUCN, Gland, Switzerland [http://www.redlist.org, accessed 13 de junio 2006).
Jara, C. (1994). Camarones dulceacuícolas en Chile. Informe técnico-científico. Instituto de Zoología, Universidad Austral de Chile.
Kruuk, H., Conroy, J. W. H. Moorhouse, A. (1987). Seasonal reproduction, mortality and food of otters (Lutra lutra L.) in Shetland. Symposium of Zoological Society of London. 58: 263-278.
Kruuk, H., Moorhouse, A. (1990). Seasonal and spatial differences in food selection by otters (Lutra lutra) in Shetland. Journal of Zoology. 221: 621-637.
Kruuk, H., Moorhouse, A. (1991). The spatial organizationof otters (Lutra lutra) in Shetland. Journal of Zoology. 224: 41-57.
Kruuk, H. (1995). Wild Otters, Predation and Populations. Oxford: Oxford University Press. 290 pp.
Kruuk, H. (2006). Otters: Ecology, Behaviour and Conservation. Oxford University Press. 265 pp.
Manly, B. F. J. (1995). Measuring selectivity from multiple choice feeding-preference experiments. Biometrics. 51: 709-715.
Mason, C. F., Macdonald, S. M. (1986). Otters: Ecology and Conservation. Cambridge: Cambridge University Press.
Medina, G. (1996). Conservation and status of Lutra provocax in Chile. Pacific Conservation Biology. 2: 414-419.
Medina, G. (1997). A comparison of the diet and distribution of southern river otter (Lutra provocax) and mink (Mustela vison) in southern Chile. Journal of Zoology. 242: 291–297.
Medina, G. (1998). Seasonal variations and changes in the diet of southern river otter in different freshwater habitats in Chile. Acta Theriologica. 43: 285-292.
Medina, G., Kaufman, V., Monsalve, R., Gómez, V. (2003). The influence of riparian vegetation, woody debris, stream morphology and human activity on the use of rivers by southern river otters Lontra provocax in Chile. Oryx. 37: 413-421.
Melquist, W. E., Hornocker, M. G. (1983). Ecology of river otters in west central Idaho. Wildlife Monographs. 83: 1-60.
Osgood, W. H. (1943). The Mammals of Chile. Publication of the Museum of Natural History, Zoological Series. 30: 1-268.
Pardini, R. (1998). Feeding ecology of the neotropical river otter Lontra longicaudis in an Atlantic Forest stream, South-eastern Brazil. Journal of Zoology. 245: 385-391.
Quadros, J., Monteiro-Filho, L.A. (2001). Diet of the neotropical otter,Lontra longicaudis, in an Atlantic Forest area, Santa Catarina State, Southern Brazil. Studies on Neotropical Fauna and Environment. 36: 15-21.
Ramírez, C., San Martín, C., Medina, R., Contreras, D. (1991). Estudios de la flora hidrófila del Santuario de la Naturaleza "Río Cruces" (Valdivia, Chile). Gayana Botánica. 48: 67-80.
Reid, D. G., Code, T.E., Reid, A. C., Herrero, S.M. (1994). Food habits of the river otter in a boreal ecosystem. Canadian Journal of Zoology. 72: 1306–1313.
Rudolph, E.(2002). Sobre la biología del camarón de río Samastacus spinifrons (Philippi 1882) (DECAPODA, PARASTACIDAE). Gayana. 66: 147-159.
Roth, J. D., Hobson, K. A. (2000). Stable carbon and nitrogen isotopic fractionation between diet and tissue of captive red fox: implications for dietary reconstruction. Canadian Journal of Zoology-Revue Canadienne de Zoologie. 78: 848-52.
Schlatter, R., Salazar, J., Villa, A., Meza, J. (1991). Demography of Black-necked swans Cygnus melancoryphus in three Chilean wetland areas. Pages 88-94 in: Sears J. & P. J. Bacon (eds.), Proceedings of the third International Swan Symposium, Oxford, England, 1989. Waterfowl, Special Supplement 1.
Somers, M. J. (2000). Foraging behaviour of Cape clawless otters (Aonyx capensis) in a marine habitat. Journal of Zoology. 252: 473-480.
Steubing, L., Ramírez, C., Alberdi, M. (1980). Energy content of water and bog plant associations in the region of Valdivia (Chile). Vegetario. 43: 153-161.
Van der Zee, D. (1981). Prey of the Cape clawless otter (Aonyx capensis) in the Tsitsikama Coastal National Park, South Africa. Journal of Zoology. 194: 467-483.
Wise, M. H., Linn, I. J., Kennedy, C. R. (1981). A comparison of the feeding biology of mink Mustela vison and otter Lutra lutra. Journal of Zoology. 195: 181-213.
Yeates, L. C., Williams, T. M., Fink, T. (2007). Diving and foraging energetics of the smallest marine mammal, the sea otter (Enhydra lutris). The Journal of Experimental Biology. 210: 1960-1970.
Zar, J. H. (1999). Biostatistical Analysis. New Jersey: Prentice Hall.
Résumé : Predation et Position Trophique de la Loutre du Chili (Lontra provocax) dans la Zone Humide «Sanctuaire De La Nature, Carlos Adwandter», Sud du Chili
Les loutres sont considérées comme des prédateurs qui participent à la régulation des écosystèmes et affectent la structure des communautés de proies en modulant leurs densités. Le régime alimentaire de la Loutre du Chili (Lontra provocax) a seulement été décrit à partir de la fréquence d'occurrence des proies dans les épreintes. A ce jour, la relation entre la disponibilité trophique et la sélection des proies par la Loutre est inconnue. Dans cette étude nous avons évalué la relation entre la sélection des proies et leur disponibilité dans différents environnements. Nous avons supposé que cette espèce est un prédateur opportuniste et que son régime alimentaire pourrait être dépendant de l'abondance en proies. Nous avons étudié les restes fécaux dans deux environnements différents (marais et rivière) dans le «sanctuaire de la nature Carlos Andwanter» au sud du Chili. Les proies retrouvées dans les épreintes ont été identifiées grâce à des collections de référence. Nous avons estimé la fréquence d'occurrence, la disponibilité relative des proies et l'indice de sélection. De plus, nous avons quantifié la disponibilité des proies à l'aide de pêches électriques. Nos résultats suggèrent que la niche trophique pour L. provocax est limitée à peu de proies avec notamment l’écrevisse (Samastacus spinifrons) comme composant alimentaire majoritaire. Quoi qu’il en soit, L. provocax consomme des écrevisses de taille spécifique indépendamment de leur disponibilité dans l'environnement. Nos résultats suggèrent que la Loutre est un prédateur opportuniste mais montre une nette sélection des écrevisses de taille moyenne.
Revenez au dessus
Resumen: Seleccion de Presa y Nicho Trófico de la Nutria de Rio Sudamericana (Lontra provocax) en Humedales del “Santuario De La Naturaleza Carlos Andwanter”, en el Sur de Chile
Las nutrias son consideradas como depredadores tope y participantes activos en el modelamiento de la estructura de las comunidades a través de la modulación de la densidad de presas. La dieta de la nutria de rio Sudamericana (Lontra provocax) ha sido descrita únicamente sobre la frecuencia de ocurrencia de las presas en la fecas. Por lo que hasta el momento se desconoce la relación entre la disponibilidad trófica y la selección de presas. En este estudio nosotros evaluamos la relación entre selección de presas y su disponibilidad en diferentes ambientes. Nosotros proponemos la hipótesis que esta especie es un depredador oportunista, y que su dieta podría estar determinada por la abundancia de las presas. Nosotros estudiamos restos de fecas en dos ambientes diferentes (hualve-rio) en el “Santuario de la naturaleza Carlos Andwanter” en el Sur de Chile. Las presas encontradas en las fecas fueron identificadas a través de colecciones de referencia. Nosotros estimamos la frecuencia de ocurrencia, disponibilidad relativa de presas, y el índice de selección. Además, nosotros cuantificamos la disponibilidad de presas usando electropesca. Nuestros resultados sugieren que el nicho trófico de L. provocax es restringido a unos pocos item presa, con el camarón de rio (Samastacus spinifrons) como el componente más importante de la dieta. Sin embargo, L. provocax consume tamaño especifico de camarones independientemente de su disponibilidad en el ambiente. Nuestros resultados sugieren que L. provocax es un depredador oportunista, pero muestra una clara selección por tamaños intermedios de S. spinifrons.
Vuelva a la tapa